Se calcula que en México actualmente existen 117 millones de hectáreas potencialmente agrícolas, de las que 23 millones son laborables
Se calcula que en México actualmente existen 117 millones de hectáreas potencialmente agrícolas, de las que 23 millones son laborables. De esta superficie laborable, 4,8 millones son de riego y el resto 18.2 millones, son de temporal (el 37 % de la superficie es temporalera regular y el 63 % es de mal temporal). De esta superficie temporalera, 10.7 millones son tierras cosechables, de las que unos 4 millones de hectáreas reciben una precipitación de – 400 mm. anuales. Unos 7.5 millones de hectáreas son improductivas, principalmente por extrema sequía1.
El problema de la resistencia de las plantas a la sequía es actual no solamente en relación a la selección de formas estables, sino también en relación al aumento de las reservas de la productividad final de las plantas. La elaboración de procedimientos que regulan el estrés hídrico en las plantas es imposible sin la consideración de sus propiedades genéticas. Como regla todos los modelos evolutivos y los procesos de mejoramiento genético se construyen en base a los cambios de los genes localizados en los cromosomas y éstos incluidos en el núcleo, por la influencia de las mutaciones o de la selección. Con esto, por completo se ignora el papel de la variabilidad de los genes del citoplasma, denominados plasmagénes, los cuales contienen ácido desoxirribonucleico (ADN) y participan en la transmisión y herencia de algunos caracteres, uno de los más importantes es la esterilidad citoplásmica masculina, de gran utilidad en la formación de híbridos de maíz, girasol, sorgo, etc.
Desde el redescubrimiento de las leyes de la herencia de Gregorio J. Mendel quedo claro que algunos tipos de variabilidad no se sujetan a estas leyes: la diferencia de los resultados en las cruzas recíprocas (cruzas directa e inversa), en la segregación de la generación F1, en la combinación independiente de los genes, etc. El análisis de las cruzas recíprocas por ejemplo de trigo (interespecíficas o intergenéricas) muestra que en los híbridos F1 existen diferencias significativas tales como: la duración del periodo vegetativo, la altura de las plantas, la resistencia de las plantas y el desarrollo de algunos elementos productivos2.
En la actualidad, las investigaciones más amplias sobre la variabilidad citoplásmica se han realizado en los híbridos alocitoplásmicos de trigo, obtenidos por el método de cruzas regresivas, en las cuales el núcleo del trigo blando Triticum aestivum L. funciona en un citoplasma de diferentes especies de Triticum, Aegilops y Secale3. En los híbridos alocitoplásmicos de trigo la dirección y la dinámica de los procesos de crecimiento y desarrollo están condicionados a las interacciones de los sistemas genéticos del núcleo (del progenitor masculino) y del citoplasma (del progenitor femenino). Las nuevas formas genéticas de trigo muestran que su utilización en el mejoramiento de plantas permitirá obtener nuevas variedades con un complejo de rasgos y caracteres valiosos4.
El objetivo de este estudio es el análisis comparativo de la productividad de una población híbrida de trigo alocitoplásmico con citoplasma Triticum timopheevi.
En calidad de objeto de investigación se utilizó el siguiente material fitogenético de trigo:
La población híbrida de trigo alocitoplásmico de primavera Moscovskaia 35 x Virovskaia 4 (con el citoplasma Triticum timopheevi Zhuk.) (Ver esquema).
La variedad testigo Enita (es de tipo intensivo, responde a la fertilización, tiene periodo vegetativo intermedio, es estable al acame, tolerante a la sequía y resistente a la roya).
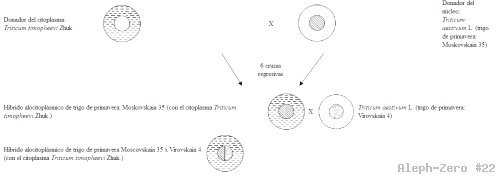
Esquema de la población híbrida de trigo alocitoplásmico de primavera Moscovskaia 35 x Virovskaia 4 (con el citoplasma Triticum timopheevi Zhuk.).
La siembra se realiza en unidades de 10 surcos de 1m. de largo, espaciados por 15 cm., la densidad de siembra es de 50 semillas por surco. Después de la cosecha se hace su análisis estructural. De acuerdo a las características morfológicas de sus espigas, se diferenciaron cuatro morfobiotipos de la población híbrida de trigo. Los resultados de algunos elementos productivos de los morfobiotipos F4 mostraron las siguientes características:
Número de granos en la espiga principal. Los morfobiotipos variaron de 31.7 +/- 2.7 (No.4) a 57.0 +/- 4.5 (No.2). En relación con enita (41.5 +/- 2.8) tuvieron especial interés los morfobiotipos No.1 (49.7 +/- 2.9) y No.2 (57.0 +/- 4.5).
Masa de granos en la espiga principal (g). Los morfobiotipos variaron de 1.60 +/- 0.1 (No.4) a 2.84 +/- 0.2 (No.2). En comparación con enita (2.51 +/- 0.1) sobresalieron los morfobiotipos No.1 (2.60 +/- 0.1) y No.2 (2.84 +/-0.2).
Masa de 1000 granos (g). Los morfobiotipos variaron de 50.2 +/- 1.8 (No.4) a 54.1 +/- 2.4 (No.1). En comparación con enita (44.9 +/- 1.5) se distinguieron todos los morfobiotipos.
Para la identificación de los morfobiotipos de trigo fue analizada la comparación de su gliadina por el método de electroforesis. Se analizaron los morfobiotipos Nos.2 y 3.
En el gel poliacrilamínico, los resultados mostraron que los morfobiotipos Nos.2 y 3 están constituidos por dos biotipos en relación: 80% - 20% y 40% - 60% respectivamente. El morfobiotipo No.2 tiene mucha semejanza entre sus biotipos componentes. Todos los biotipos tienen idénticas las b- y g-zonas, / b-23145 / y / g-1 34 / respectivamente.
En el gel de almidón, los resultados señalaron que los morfobiotipos Nos.2 y 3 poseen polimorfismo en la composición de su gliadina y están constituidos por dos biotipos en relación: 90% - 10% y 70% - 30% respectivamente. Entre los biotipos identificados, los más perspectivos para la selección de variedades con calidad de grano, tienen fórmula genética de gliadina 14.1.5.1.1.1., el polimorfismo aumenta la diversidad genética y ofrece grandes posibilidades para obtener el biotipo con las características deseadas.
En base al análisis de las investigaciones se puede concluir que fueron diferenciaron nuevos morfobiotipos de trigo con citoplasma Triticum timopheevi, cuya particularidad es la presencia de gran masa de 1000 granos. Dichos morfobiotipos son formas genéticas heterogéneas de trigo. En la selección de variedades de trigo con calidad de grano, se puede utilizar en cada morfobiotipo el segundo de sus biotipos componentes.
- López T., M., Resistencia de las plantas, ESAHE, Trillas, México, 1995.
-
Spetsov, P. and Tsunewaki, K., “Increasing genetic variability in common wheat by utilizing alien cytoplasms”, Jap. J. of Breed, 36(3): 263-271, 1986.
-
Beavis, W.D., Pollak, K.E. and Frey, J.A., “Theoretical model for quantitatively inherited traits influenced by nucleo – cytoplasmic interactions”, Theory and Applied Genetics, 74(4): 571–578, 1987.
-
Reyes M., J.M., “Selección de trigo alocitoplásmico de primavera con Triticum timopheevi Zhuk. citoplasma”, XVII Congreso de Fitogenética, Acapulco, México, p. 383, 1998.